Previous: Computer graphics and computer vision color models Up: Color Models
Next: De Valois & De Valois's color model
Probably the best known opponent model of color perception is the
opponent process theory of Hurvich and Jameson, described in a series of
articles
([Jameson \& Hurvich 1968][Hurvich \& Jameson 1956][Jameson \& Hurvich 1956][Hurvich \& Jameson 1955][Jameson \& Hurvich 1955],
see also [Hurvich 1981]). The roots of this theory can be traced
back to Hering's theory of color vision [Hering 1878] in terms of
opponent hues which cancel each other when
superimposed:
yellow and blue on the one hand, and green and red on the other. Hurvich
and Jameson developed an experimental procedure which they called hue
cancelation, which allowed them to quantitatively express the relative
amounts of each of the four basic hues present in any spectral stimulus.
Figure
represents an example of the resulting
chromatic response or chromatic valence functions.
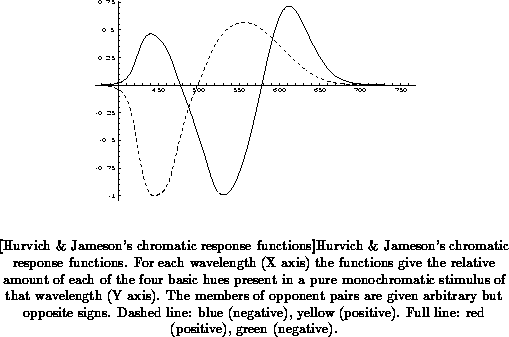
The chromatic response functions are determined experimentally for each
individual observer, but, although there are slight interpersonal
differences, the general shape of these functions remains the same. The
functions presented in Figure are derived from the
CIE 1931 standard observer color matching functions, and may be taken as
representative for a large number of observers. Chromatic response
functions are assumed to correlate directly with hue perception. In
addition to the chromatic response functions, an achromatic response
function is determined, which represents the perceived brightness of a
stimulus of a given spectral composition. Based on the chromatic response
functions, Hurvich and Jameson also derived hue coefficient functions
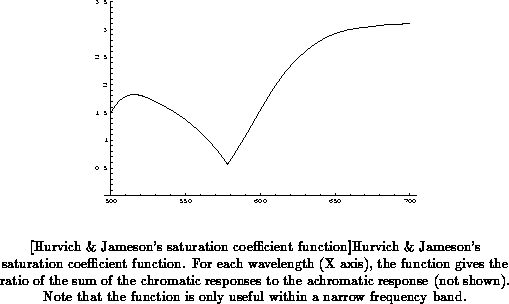
(Figure
) and saturation coefficient functions
(Figure
).
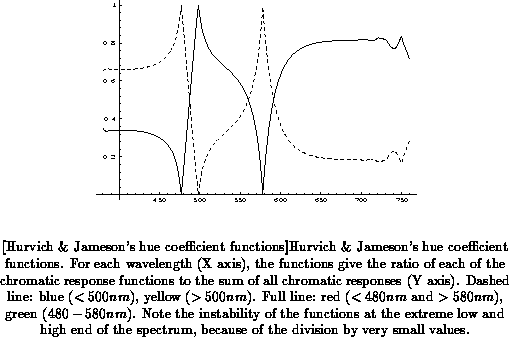
The hue coefficient functions express hue as the ratio of each chromatic
response to the sum of all chromatic responses, at each wavelength. Thus,
each of the four basic or physiologically primary hues has associated
with it a function which varies between 0 and 1 over the visible wavelength
spectrum. The wavelengths where these functions reach 1 are the loci of the
unique or pure hues. Only unique blue, green, and yellow exist
as spectral colors, but not red, as shown in Figure . The
saturation coefficient functions give the ratio of the sum of the chromatic
responses to the achromatic response at each wavelength, and are supposed
to reflect the perceptual saturation of monochromatic stimuli of different
wavelengths. Hurvich and Jameson showed that their model could account well
for the stimulus attributes of spectral lightness, hue, and saturation, and
their associated psycho-physical functions. They also showed how their
model could account for some changes in perceived brightness, saturation,
and hue with chromatic adaptation, and to some extent for color constancy
phenomena. They finally defined a ``psychological color specification
system'', which is a hue/saturation/brightness (HSB) color space based on
hue and saturation coefficients in a polar coordinate system, and
achromatic response as the brightness dimension.
Hurvich and Jameson's model is the first quantitative color opponent model based on psychophysical experimentation, and as such deserves a lot of credit. They demonstrate that an impressive range of psychophysical phenomena can be explained elegantly in terms of their model. However, the model has a number of drawbacks for use as a computational model of color perception and color naming, which I now outline briefly. One could interpret the hue coefficient functions as categorial perception functions, but it is not clear how to extend these to colors other than the four physiologically unique hues (as Hurvich and Jameson call them). All their data deal only with some (moderate) luminance levels, and there seems to be no systematic way to extend it to arbitrary luminance and adaptation levels. Their discussion of adaptation phenomena is also limited to some specific steady-state adaptation states, which require the spectral composition of the adapting light to be known beforehand; i.e., the model is not useful as a computational model of color perception if it is to operate in real-world environments where lighting characteristics are not known. In addition, their model says nothing about the dynamics of adaptation with respect to time. With the requirements for computational models of color perception in mind, Hurvich and Jameson's model is to some extent a descriptive rather than an explanatory model. Another problem with their discussion of adaptation is that it implicitly assumes that the different photoreceptor types (the cones in the retina with different spectral sensitivities) can change their adaptation states independently of each other. In view of their assumption that adaptation is proportional to receptor output, and the large degree of overlap of the spectral sensitivities of the photoreceptor types they propose (or even the ones currently accepted), this independence is very unlikely. [Wyszecki \& Stiles 1982] discuss problems with another assumption underlying Hurvich and Jameson's work, viz., the linearity laws for opponent hue cancelation. These laws seem to hold for the red/green responses, but not under all circumstances for the yellow/blue responses. The color space defined by Hurvich and Jameson is only partial; there is no representation of an actual 3D color solid, only selected planar sections through it at fixed luminance levels. The regular spacing of hues in the polar coordinate system (red diametrically opposite green, and yellow to blue, with the axes perpendicular to each other) is unlike that of, e.g., the Munsell color model. As I mentioned above, the categorial boundaries of the four basic hues (red, green, yellow, and blue) are ``built in'' to the color space, and it is not clear how other perceptual color categories would fit in. The basic organization of my own color space using opponent color pairs is of course similar to that of Hurvich and Jameson.
When compared to neurophysiological data, particularly [De Valois et al. 1966], the red response in the short wavelength region is strikingly absent, although the overall similarity is clear. It is not so clear how this discrepancy can be explained, other than to postulate some unknown higher level neural mechanism that would be responsible.