Previous: Saturation levels revisited Up: Modeling Neurophysiological Data
Next: The sigmoid-of-linear activation model
As discussed in Section (p.
),
[De Valois \& De Valois 1975] suggest that the six types of LGN cells found can
be grouped into three dimensions, when mirror image pairs
are combined into one
dimension. That results in a red-green, a blue-yellow, and a brightness
dimension which can be arranged in a ``double cone''-type color space. We
now turn to the construction of such a 3-dimensional color space based on
the six response functions described above. In view of the hypothesis that
mirror-image coding of signals in the nervous system is a way to increase
the dynamic range of the signal
[Slaughter 1990], and following the suggestion of
[De Valois \& De Valois 1975], I will assume that the responses of mirror-image
pairs of functions can be added to give one composite function:
where is a new composite green--red opponent function,
a new
composite blue--yellow opponent function,
a new composite brightness
function (non-opponent), and
represent the six response
functions
,
,
,
,
, and
, respectively. The
members of the pairs have to be subtracted rather than added because they
are 180 degrees out of phase relative to each other, and we want the
corresponding phases to add up rather than cancel each other. The order of
the terms determines the sign of the phases of the composite functions, and
is arbitrary. I will always use the order of
equations
--
as the
convention. Figure
(upper half) shows the resulting
opponent functions at a relative radiance of 0.5.
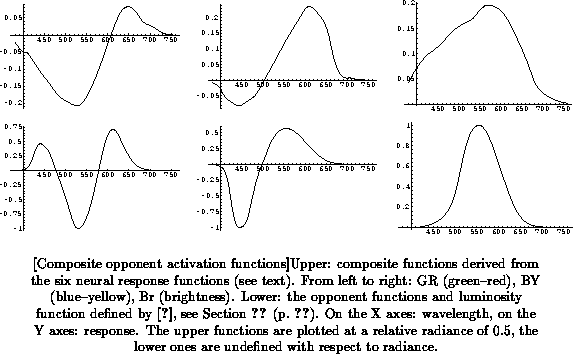
Some interesting properties of these functions are:
In my opinion, these observations lend support to the assumption of one
global maximum firing rate, and to the method of combining the six
component functions into three. I therefore feel confident in using the
three composite functions obtained as a neurophysiological basis for a
3-dimensional color space with a color opponent organization, interpreting
the and
functions as color opponent dimensions and the
function as the brightness dimension. Later I will investigate the
usefulness of this color space for the color naming problem. For now, I
would like to note that this approach may provide an interesting bridge
between the neurophysiology and the psychology of color perception (see
Section
, p.
).